
A géntechnológia tudományos eredményei és társadalmi hatásai
Új korszak a növénybiológiában és -nemesítésben
Dudits Dénes
A tudomány eddigi története számos példával szolgált arra, hogy egy-egy zseniális felismerés nyomán a kutatás alapjaiban új metodikai háttere jött létre, ami aztán lényeges szemléletbeli változásokhoz is vezetett. Ilyen általános hatású, nagy horderejű eredménynek tekinthetjük a rekombináns DNS-módszerek kidolgozását, és ez a biológiai tudományok szinte minden területén jelentős, új felismerések megszületését tette lehetővé. A 70-es években vált lehetségessé a DNS-molekulák in vitro módosítása: restrikciós endonukleázokkal történő feldarabolása, ligázokkal való összekapcsolása, kör alakú plazmid DNS-ek segítségével adott szekvenciarészek felszaporítása (klónozása). Elkészültek az első génkönyvtárak, megvalósíthatóvá vált a gének izolálása. A nukleotidok sorrendjének meghatározására kidolgozták a DNS-szekvenálás módszerét. A DNS kémiai szintézise szintén fontos eleme a rekombináns DNS-technológiának. A rövid DNS-szakaszok szintetizálása különösen nélkülözhetetlenné vált a polimeráz láncreakcióval (PCR) megvalósított DNS- molekula megsokszorosításoknál. A kémcsőben kialakított DNS-molekulák a transzformáció módszerével visszaépíthetők a sejtekbe, biztosítható az idegen, transzgén működőképessége. Új, hírvivő RNS-molekulák, fehérjék szintézisével megváltoztathatók a sejtek funkciói, új öröklődő tulajdonságok alakíthatók ki.
Teljesen magától értetődő, hogy a mikrobiális kísérleti rendszerekben kidolgozott megközelítések a növénybiológusok számára is izgalmas kutatási lehetőségeket kínáltak. Az első növényi génizolálási próbálkozások eredményeiről 1979-ben Edinburghban megrendezett konferencián számoltak be a kutatók. Ekkorra már idehaza is folytak növényi DNS klónozására épülő kísérletek. Az MTA Szegedi Biológiai Központ Genetikai Intézetében Koncz Csabának és munkatársainak a kukorica mitokondriumában található plazmid DNS klónozásáról készült közleménye 1981-ben jelent meg. Ez jelentette a növényi géntechnológia hazai kezdetét [1]. A legfontosabb módszereket ez ideig csak kevés magyar nyelvű könyv ismertette [2, 3, 4]. A géntechnológia és a kapcsolódó molekuláris módszerek az elmúlt 20 év alatt nemcsak a növények életjelenségeinek megismerésében váltak nélkülözhetetlenné, hanem a fajtaelőállító növénynemesítés eszköztárát is sokban gazdagították. Az alábbiakban néhány példa segítségével mutatjuk be a rekombináns DNS-módszerek használatára épülő megközelítések lényegét, különös tekintettel a nemesítési felhasználásra. A génizolálás és - beépítés alapot ad a növények agronómiai hasznosíthatóságát javító tulajdonságok kialakításához. Így egyre intenzívebben folyik a géntechnológia módszereit felhasználó növénynemesítés. Köztermesztésben már vannak fajták, amelyeket ezen az úton állítottak elő. A géntechnológiával nemesített (GN) fajták (a sajtóban genetikailag módosított szervezeteknek, GMO-nak nevezik ezeket a növényeket) terjedésével kapcsolatban igen heves viták alakultak ki az előnyöket és kockázatokat illetően. Ezért indokolt, hogy - néhány szempont felvázolásával - kísérletet tegyünk az álláspontok közelítésére.
A DNS-szekvenciától a funkcióig: növényi genomprogramok
A DNS-molekula nukleotid-sorrendjében tárolt információ részint az RNS- és fehérjemolekulák szintézisének (transzkripció, illetve transzláció) irányításán keresztül meghatározza a sejtekben lejátszódó folyamatokat, részint kromoszómákba való szerveződést követően az információ átkerül az osztódással létrejött utódsejtekbe, így biztosítva az információ megőrződését, öröklődését. A géneket, az örökítő anyag funkcionális egységeit a genetikusok hosszú ideig csak a felszínen megjelenő tulajdonságok, a fenotípus vizsgálatával tudták azonosítani és tanulmányozni. A rekombináns DNS-módszerek kidolgozásával vált lehetővé a gének izolálása, szerkezeti elemeinek behatárolása és módosítása. Sor kerülhetett nagyszámú növényi gén szekvenciájának meghatározására, majd pedig felismerhetővé váltak a legfontosabb funkcionális elemek, mint ahogy azt az 1. ábra bemutatja. Az 5' végen elhelyezkedő ún. promoterszakasz felelős a gén működésének szabályozásáért. Az itt található szekvenciamotívumok (cisz-elemek) specifikus fehérjéket (transzkripciós faktorokat) képesek megkötni. Az így kialakuló fehérje-DNS kölcsönhatás felelős lehet az mRNS (messenger [m], hírvivő RNS, l. 1. ábra) szintézisének megindításáért vagy a gén működésének leállításáért. A növény életciklusának különböző szakaszaiban a gyökérben, a szárban vagy a virágban más-más gének működnek és irányítják a sejtek funkcióit, a szervek differenciálódását. A gének promoterei döntő szerepet játszanak a szövet- vagy szervfüggő génműködés biztosításában. Vannak olyan promoterek, amelyek csak akkor kapcsolnak be, ha a növényeket valamilyen stresszhatás éri. Természetesen azonosíthatók alapvető, minden sejt működéséhez szükséges fehérjék, enzimek is. Az ezeket meghatározó gének promoterei folyamatos, ún. konstitutív működést mutatnak. A biotechnológiai alkalmazások szempontjából kiemelt jelentősége van a promoterként funkcionáló DNS-molekuláknak, hiszen segítségükkel érhető el a célnak megfelelően szabályozott génműködés. Ezért külön géntechnológiai programok szolgálják a promoterek izolálását; az így kapott szekvenciák gyakran szabadalmi védelmet kapnak, és eladható termékként jelennek meg.
A növényi gének is rendelkeznek az ún. kódoló régióval, amelynek DNS-szekvenciája először hírvivő RNS-sé íródik át, majd megtörténik a fehérje szintézise. A gén 3' végén található még az ún. terminációs jel, amely behatárolja az mRNS-molekula hosszát. A génizolálás célja sok esetben nem az ún. genomikus klón megkeresése, hanem az mRNS-ről készült kópia-DNS (cDNS) in vitro szintézise. A cDNS-ek reverz transzkripcióval nyerhetők, amelyek a sejtekben éppen működésben lévő géneket reprezentálják. Mind alapkutatási, mind alkalmazási szempontból figyelemre méltó információkhoz juthatunk, ha különböző állapotú sejtekből származó mRNS- molekulákról készítünk cDNS-klóntárakat, valamint azokban a megegyező és eltérő cDNS-klónokat azonosítjuk. Így például összevethető a baktérium által fertőzött, illetve nem fertőzött, a hidegkárosított és normál hőmérsékleten nevelt gyökér-, levél-, szár eredetű sejtekben a génkifejeződési mintázat, és felismerhetők az egyes sejtállapotokra jellemző cDNS-klónok. A differenciális génkifejeződés tanulmányozásában az utóbbi években jelentős előrehaladást jelentett a DNS-chip technológia* megszületése. Nagyszámú, akár több tízezer cDNS-klónt - egy szigorúan meghatározott rendszerben, robot segítségével - üvegfelületre rögzítenek, amihez fluoreszcens próbák hibridizálhatók, különböző eredetű sejtekből származó minták felhasználásával. A hibridizációs jel erősségének mérését, valamint a specifikus fluoreszcens jelek szétválasztását lézerleolvasó készülék végzi. A digitalizált jelek analízisével azonosíthatók azok a cDNS- klónok, illetve az általuk képviselt gének, amelyek meghatározott sejtállapot esetén vagy erős kifejeződést mutatnak, vagy kikapcsolt állapotban vannak. A cDNS-klónok szekvenciájának ismeretében a nemzetközi adatbázisok segítségével megállapítható, hogy az adott gén egy ismert vagy ismeretlen fehérjét kódol-e. Ha új, az adatbázisban nem szereplő fehérjét azonosított a vizsgálat, már akkor is - a kifejeződési mintázat alapján - bizonyos funkcionális információhoz juthatunk. Részletes felbontású transzkripciósmintázat-értékelés megbízható kiindulópontot jelent a funkciók behatárolásához.
Több növényfaj bevonásával is folyamatban van különböző eredetű génkönyvtárakból véletlenszerűen kiválasztott cDNS-klónok szekvenciájának meghatározása. A nagy teljesítményű, automata DNS-szekvenátorok egy-egy növény esetében akár több tízezer cDNS-t mint génkifejeződési markert (expressed sequence tag, EST) is azonosíthatnak. Napjainkban a legnagyobb EST adatbázisok rizs és Arabidopsis** esetében állnak rendelkezésre. Az EST-klónok 25%-a már ismert gén megfelelője. Az EST cDNS-ek igen fontos szerephez jutnak a genetikai és molekuláris térképek elkészítésénél. Az EST programokkal párhuzamosan folyik nagyméretű (300-1000 kb [ahol 1 kb = ezer nukleotid]) növényi genomikus DNS-szakaszok klónozása bakteriális vagy élesztő vektorokba (bacterial artificial chromosome, BAC; yeast artificial chromosome, YAC). Az ilyen génkönyvtárak jelentik az alapját a sokszor nemzetközi összefogással megvalósuló szekvenálási programoknak, amelyek célja valamely növény teljes DNS- állományára kiterjedően a nukleotidsorrend megfejtése. A legfrissebb közlemények szerint az Arabidopsis 2. és 4. kromoszómájának megszekvenálása sikeresen befejeződött [5, 6]. Ez fontos mérföldkő a teljes genom DNS-szekvenciájának meghatározását célzó programokban. Feltételezhető, hogy a nukleotidsorrend megfejtése sikeresen halad a rizs és kukorica esetében is. Két kromoszóma adatai alapján 40-60% az ismeretlen gének aránya. Érdekes eredmény, hogy igen nagymértékű genomátrendeződésre utaló szekvenciamotívumok találhatók. Magas a gének, illetve a kromoszómarégiók megduplázódásának gyakorisága.
A folyamatban lévő szekvenálási programok jelentős szellemi és anyagi hátteret igényelnek. Szinte elképzelhetetlen, hogy a felszínre került információkat ne használják fel a növénybiológiai alapkutatásokban, illetve az alkalmazott molekuláris növénynemesítésben. Különösen így lesz, ha minél több szekvenciarészlet esetében megállapítható a kódolt fehérje szerepe is akár szekvencia-összehasonlítás, akár fehérje-fehérje kölcsönhatás segítségével. Az ún. funkcionális genomprogramok többféle megközelítést is felhasználnak. Az evolúció során struktúrájukban és funkciójukban konzervált fehérjék jellemzése megtörténhet heterológ sejtekben. Így élesztőmutánsok eredményesen használhatók növényi cDNS-ek funkciójának megállapításához. Mutációk kijavítása, komplementációja vezetett például több növényi sejtciklusgén izolálásához. Igen kiterjedten használják az élesztő kéthibridrendszert fehérje-fehérje kölcsönhatásban résztvevő partnerek génjeinek klónozására. Ebben a kísérleti rendszerben az élesztősejtek akkor szaporodnak, ha két növényi cDNS-t hordozó plazmiddal történt transzformáció után a szintetizálódó fehérjék komplexeket képeznek. A kölcsönhatás ténye már funkcióra vonatkozó információ, amelyet természetesen más módszerekkel is indokolt megerősíteni. A gén és funkció összekapcsolásában kiemelt szerephez jutnak az inszerciósmutáns-gyűjtemények. Ezekhez úgy juthatunk, hogy ugráló gének, ún. transzpozonok vagy ismert szekvenciájú idegen DNS-szakaszok beépülnek a növényi gén bármely régiójába, és így a gén elveszti működőképességét. A gén működési hibája folytán megváltozott mutáns fenotípus alakul ki. Mivel a mutációt kiváltó DNS szekvenciája ismert, a beépülésük helyét közrefogó DNS-szakasz, az elrontott gén izolálható és szekvenciája meghatározható.
Ezzel kezdetét veszi a gén funkcionális analízise, amikor kapcsolat teremthető egy fenotípus és a szekvenciaadat között. A gének funkciójára vonatkozó közvetlen információt a génbeépítés, az ún. transzformáció módszerével nyerhetünk. Ha működőképes formában akár genomikus klón promoterrégióját vagy kódoló szekvenciáját, akár cDNS-t visszajuttatunk növényi sejtekbe, melyekből aztán transzgenikus növényt nevelünk fel, akkor a fenotípusos bélyegek megváltozásából következtetni lehet a gén szerepére. A transzformáció módszere elengedhetetlen eleme a funkcionális genomprogramoknak. Magától értetődő törekvés, hogy ha a növénybe beépített gén kifejeződésének eredményeként agronómiai szempontból kedvező változás következik be, akkor azt a növénynemesítő hasznosítani akarja. Igen visszás helyzetet szülne, ha az így nyert értékes tenyészanyagokat csak azért kellene kidobni, mert azok izolált gén beépítése nyomán alakultak ki. Mint korábban láthattuk, a szekvenálási programok a teljes DNS-állományt veszik célba. Ezzel egy időben alig találhatunk növényi szervet, életjelenséget, amellyel kapcsolatban ne lenne folyamatban génizolálási munka és természetesen transzformánsok előállítása. Könnyen belátható, hogy ezek közül azok a géntechnológiai megoldások, amelyek nemesítik a növényeket, azaz emberi igényeket kielégítő terméket biztosítanak, előbb vagy utóbb felhasználásra kerülnek. Nem véletlen, hogy a legutóbbi években ugrásszerűen megnőtt a transzgenikus technológiára alapozott szabadalmak száma. Ennek az igen intenzív technológiai fejlesztésnek az alapját a legújabb tudományos eredmények jelentik, így volt ez a növénynemesítés korábbi története során is. Gondoljunk a citológiai megfigyelésekre épülő poliploid nemesítés sikereire. Néhány évtizede elegendő információt jelentett a kromoszómák számának meghatározása. Napjainkban ez már kevés, és a módszerek a genetikai anyag mélyebb szerveződési szintjeinek feltárására hivatottak. Mindinkább a sejtfunkciók, a fejlődési program genetikai szabályozásának központi problémájává válik annak tisztázása, hogy a sejtet ért külső jelek miként tevődnek át a transzkripciós változások szintjére. Az egyes jelátviteli láncolatok koordinációjára épül az egyedfejlődés belső programozottsága, a külső környezeti tényezőkre adandó válaszreakciók létrejötte. A génizolálási kísérletek megközelíthetővé teszik ezeket az alapvető biológiai kérdéseket, ugyanakkor hozzájárulhatnak a növénynemesítés eredményességéhez is.
A transzgenikus növények előállításának módszerei
A nemzetközi gyakorlatban és így automatikusan a hazai sajtóban is, a génbeépítéssel létrehozott transzformánsokat genetikailag módosított szervezeteknek (genetically modified organisms, GMOs) nevezik. Ez a nem igazán szerencsés szóhasználat sok félreértésre ad lehetőséget. Nem szabad figyelmen kívül hagyni azt a tényt, hogy minden köztermesztésben lévő gazdasági növényünk genetikailag módosított szervezet, hiszen amikor a nemesítő keresztezést végez és szelektálja a kívánt tulajdonságú növényeket, akkor drasztikusan megváltoztatja a növény génállományát. A transzgenikus növények előállítása csak módszerében új nemesítési eljárás, de a lényeg ebben az esetben is új génkombinációk létrejötte. Indokoltabb lenne transzgenikus növényekről, illetve géntechnológiával nemesített (GN) fajtákról beszélni. Természetesen vannak lényeges megkülönböztető jegyei is ennek a technológiának. Elsőként említhetjük, hogy míg a nemesítői munka során az ivarsejtek, illetve a megtermékenyített petesejt, a zigóta közvetíti az új génvariánsok létrejöttét, addig az izolált géneket, a mai gyakorlat szerint, testi, ún. szomatikus sejtekbe építjük be. Ennek megfelelően külön feladatot jelent fertilis növények felnevelése a transzformált sejtekből. A növények esetében az egyedfejlődési program rugalmassága folytán a differenciált levél- vagy gyökér eredetű sejtekben is kialakulhat totipotens állapot, és embriogenezisen vagy organogenezisen keresztül teljes értékű, földbe kiültethető növény nyerhető egyetlen tenyésztett sejtből. Napjainkra szinte valamennyi termesztett növény szövettenyészeteiben kidolgozott a növényregeneráció metodikája. A génbeépítés másik fontos eleme a transzformációs vektor, amelynek főbb alkotóit az 2. ábra szemlélteti. A plazmid DNS-molekula több funkcionális részt is tartalmaz. Transzformálás során csak néhány sejtbe kerül be az idegen DNS, és még ritkább a sejtmagi vagy organellum DNS-be történő integráció bekövetkezése. A transzformált sejtek, növények kiválasztását a rezisztenciamarkert hordozó szelekciós blokk biztosítja. Az itt elhelyezkedő antibiotikum- vagy herbicidrezisztencia-gént valamilyen konstitutív promoter működteti. Az utóbbi időkben megfogalmazott fenntartások miatt az antibiotikumrezisztencia-gének használata háttérbe szorul, és növényi eredetű markerek kapnak elsőbbséget. A biológiai hatást biztosító gén lehet cDNS vagy genomikus DNS-szakasz. A hatékony termék-előállítás érdekében indokolt lehet mesterségesen szintetizált gén beépítése a vektormolekulába. Ilyenkor növényi kód használatával növelhető a fehérjetermék mennyisége.
A transzformációhoz használt gén igen gyakran növényből származik, sok esetben ugyanannak a fajnak a génjét építik vissza. A promoter megváltoztatása, irányított mutációk kialakítása a meglévő anyagcsere-folyamatok lényeges módosítására, új tulajdonságok kialakítására ad lehetőséget. Különösen eredményes lehet az ún. antiszensz megközelítés, amikor a gén kódoló régióját fordított orientációban építik össze a promoterrel, és a képződő antiszensz RNS- molekulák hibridizációval hatástalanítják a növény eredeti génjéről szintetizálódott mRNS-molekulákat. Ennek következtében nem keletkezik fehérjetermék, elveszik egy funkció. Ezt a stratégiát sikeresen használták például a paradicsom érésének befolyásolására. Az etilén érést serkentő növényi hormon, szintézisét gátolva piacképes terméket lehetett forgalomba hozni. A növényi DNS-szekvenciák felhasználásával megvalósított transzformáció elvben nem különbözik a keresztezéssel történő génátviteltől. Integráció után a beépített gén a recipiens növény többi génjéhez hasonlóan viselkedik. Az eddigi tapasztalatok azt mutatják, hogy a mendeli törvények szerint öröklődnek, ki vannak téve molekuláris módosításoknak, mint pl. a metilációnak. A növényi géneket felhasználó géntechnológiai eljárások esetén kevésbé kell számolni nem várt mellékhatásokkal. Egyetlen izolált gén beépítése nagyfokú specifitást, a hatások pontosabb irányíthatóságát teszi lehetővé. Keresztezést követően nagyszámú gén véletlenszerű kombinálódása következik be. Ezért a növénynemesítő hasadó utódgenerációkban végzi el a szigorú szelekciót. Eltávolítja a nem kívánt tulajdonságokat hordozó egyedeket, további szaporításra és vizsgálatra csak az értékes tenyészanyagot használja. Növények esetében a transzformáció jelenlegi módszerei nem teszik lehetővé az idegen gén integrációjának irányítását. Nem működnek hatékonyan a homológ rekombinációhoz szükséges mechanizmusok, így a beépülés véletlenszerű. Gén integrálódhat funkcionálisan fontos DNS-szakaszba, ami inszerciós mutációt eredményezhet. A környező DNS-régiók is befolyásolhatják a beépített gén kifejeződését. Bekövetkezhet a gén elhallgattatása vagy kontrollálatlan működése. Sokszor a beépített DNS egyszerre több példányban is megtalálható a transzformánsok genomjában. Ezek a bizonytalan események azonban nem akadályozzák meg használható tenyészanyagok előállítását. A keresztezéses nemesítés gyakorlatához hasonlóan több generáción át kell értékelni a transzformációból származó növények agronómiai és piaci értékeit, és csak a kívánt tulajdonságokkal rendelkező növények maradnak bent a nemesítési programban. Mindezek szükségessé teszik akár több száz független transzformáns előállítását és a több generáción át folytatott szelekciót, mielőtt sor kerül a fajtaminősítésre és a GN növény termesztésére. A transzformációs módszer a növénynemesítés integráns része, és csak az alapanyag-előállítás fázisában különbözik a többi nemesítési eljárástól.
A transzformációs vektor kialakításakor növényi promoterrel összeépíthetők vírusból, baktériumból, élesztőből vagy akár állatokból származó DNS-szakaszok. Számos példa ismert arra, hogy ezek a heterológ gének kifejeztethetők transzgenikus növényekben, ami hasznos tulajdonságok megjelenéséhez vezethet. A genetikai beavatkozásnak ezen esetei már alapvetően eltérnek a keresztezéses nemesítés során lejátszódó genetikai rekombinációs folyamatoktól. A génbeépítés következtében a korábbiakban nem létező génkombinációk jönnek létre, új anyagcsereutak és termékek alakulhatnak ki a növényekben. Mivel megnő az előre nem jelezhető hatások valószínűsége, ezekben az esetekben szükségessé válhat a transzformánsok még sokoldalúbb értékelése. Gén- és növénykombinációnként változhat a transzformáció hatása, ami kizárja, hogy általános érvényű következtetéseket vonjunk le akár sikeres, akár sikertelen kísérletekből. A több tízmillió hektáron folytatott termesztési tapasztalat arra utal, hogy agronómiai értékű fajták állíthatók elő virális vagy bakteriális gének növényekbe történő beépítésével. Ez ideig nem tudunk biológiai katasztrófáról a géntechnológiával nemesített fajták esetén.
A géntechnológiára támaszkodó nemesítés eredményessége érdekében kiemelt figyelmet célszerű fordítani a promoter eredetére és működésének sajátosságaira. A promoter kiválasztásakor a kialakítandó tulajdonság, az előállítandó termék alapvetően körülhatárolja a lehetőségeket. Biológiai aktivitással rendelkező anyagok termeltetésére a növények mint biofermentorok előnyösen hasznosíthatók. Gazdaságos hatóanyag- előállításhoz célszerű az idegen gént egy magas kifejeződést biztosító, konstitutív promoterrel működtetni a transzformáns növényekben, amelyek aztán ipari felhasználásra kerülnek. Számos géntechnológiai megoldásnál indokolatlan a beépített gént folyamatosan kifejeztetni, mert a gén termékére csak bizonyos szervekben, szövetekben, a fejlődés egy rövid szakaszán van szükség. Felesleges például egy gyomirtószer-rezisztenciát eredményező gént a szója vagy kukorica termésében aktív állapotban tartani. A rezisztenciagén által kódolt fehérjének a vegetációs periódus korai szakaszán van szerepe, amikor a gyomirtó szert kipermetezik. A későbbiekben a virágzási és magkötési stádiumban a terület borítottsága gátolja a gyomosodást. A génizolálási programok feltárnak olyan géneket, amelyek a zöld vegetatív szövetekben fejeződnek ki, az ivarszervekben és a fejlődő magban nem működnek. Ezek promotereivel biztosítható, hogy a fogyasztásra kerülő termék mentes legyen a rezisztenciagén termékétől. A géntechnológiai beavatkozások precizitásának növelésével a vásárlói fenntartások mérsékelhetők, hiszen megoldható, hogy a transzformációval előállított fajta termése azonos összetételű legyen a kiindulási fajtáéval.

A virális CaMV35S promoter fő funkcionális elemei és működésük jellegzetességei a ß- glukuronizád riportergén működése alapján. A TATA boxot és az AS-1 cisz elemeket tartalmazó A domén az embrió gyökérkezdeményében valamint a hajtás- és gyökérmerisztémában biztosít aktivitást. A teljes hosszúságú B elem a sziklalevelekben (mag és csíranövény), illetve a kifejlett növény valamennyi szövetében működteti a riportergént
Az utóbbi hónapok sajtóvitáiban kiemelt figyelmet kapott a karfiol mozaik vírusból származó ún. CaMV35S promoter. Mint a 3. ábrán [7] látható, ez a 343 nukleotidból álló DNS- szakasz több transzaktiváló faktort tud megkötni a növényi sejtekben, és sajátos génkifejeződési mintázatot eredményez a transzformáns növényekben. Ezt a konstitutív promotert igen széleskörűen használták transzgenikus növények előállításakor. A kísérleti értékelés alatt lévő tenyészanyagok, illetve termesztésbe vont fajták többsége hordozza ezt a rövid DNS-szakaszt. Annak ellenére, hogy nem állnak kísérleti adatok rendelkezésre, felmerült, hogy ez a szekvencia részt vesz nagymértékű genomátrendeződésekben és horizontális génátvitel esetén az emberi egészséget is veszélyeztetheti [8].

A polipirimidinekben (timin, citozin) gazdag régiókat tartalmazó konstitutív lucerna H3 hiszton gén promotere magas ripotergén-kifejeződését biztosít dohány transzformánsokban. ß-glukuronizád (GUS) enzim aktivitása 4-metilumbelliferil- glukuronid szubsztrát jelenlétében
Az indulatoktól fűtött vitákban nehéz eligazodni, különösen olyan kérdésben, amelyet kísérletesen nem vizsgáltak. Sürgetve az ilyen kutatások elindítását, néhány szempont felvetése talán segít a véleményalkotásban. A növényi vírusok kutatói szerint Angliában a piacon árult karfiol és karalábé 10%-a fertőzött ezzel a vírussal. Ez azzal jár, hogy a növény minden egyes sejtjében 100.000 kópiában van jelen a teljes vírus DNS. Ezzel szemben a transzformáns növények sejtjeiben 1-5 példányban található a 343 nukleotidnyi DNS-darab. Mindez azt jelenti, hogy tízezerszer több vírus- és promoterszekvenciát fogyasztanak az emberek, mint amennyi a transzformáns növényekben megtalálható. Ezeket az adatokat vette figyelembe az amerikai mezőgazdasági minisztérium, amikor évekkel ezelőtt a kockázatot elhanyagolhatónak minősítette. Érdemes megemlíteni, hogy a CaMV35S promotert hordozó transzformánsokkal végzett etetési kísérletek nem mutattak káros elváltozást csirkében, halakban, szarvasmarhában és patkányban [9]. Addig is, amíg kísérleti eredmények birtokában a kérdés megnyugtatóan megválaszolható lesz, indokolt alternatív promoterek után kutatni. A növényi eredetű regulátor szekvenciák közül jelentősége lehet a lucernából izolált hisztonpromoternek. Mint az a 4. ábrán látható, ez a promoter erősebb kifejeződést biztosít, mint a CaMV35S promoter. Hasonlóan jól használhatók a növényi aktin vagy ubiquitin gének promoterei is. Az 1. színes kép egy olyan növényi promoter működését teszi szemléletessé, amelynek 284 nukleotidból álló szabályozó régiója elegendő ahhoz, hogy egy riporten gént merisztémaszövetekben működtessen. A kék indigó festék megjelenése jelzi azokat a szövetrészeket, amelyekben a promoter funkcionál. Várható, hogy a növényi eredetű promoterek választékának bővülésével a géntechnológiával nemesített fajták elterjedése felgyorsul.


A felhasználás céljának megfelelően a rekombináns DNS-módszerekkel kialakított transzformációs vektor, mint egy kör alakú DNS-molekula, különböző módszerekkel juttatható be a növényi sejtekbe. A legkiterjedtebben használt génbeépítési megközelítés egy természetes génátviteli mechanizmusra épül. Az Agrobacterium talajbaktériumok Ti-plazmidján megtalálhatók azok a funkcionális elemek, amelyek szükségesek ahhoz, hogy sebzett növényi részekkel érintkezve egy 23 kb nagyságú DNS-szakasz (T-DNS) kivágódjék, majd a sejtmagi DNS-be integrálódjon. A beépülésben kitüntetett szerepe van az ún. határszekvenciáknak, amelyek közé elhelyezett bármely DNS-szekvencia a növényi szövetek Agrobacteriummal történő fertőzésével a növényi genomba építhető. A 2. színes kép transzformáns burgonyahajtás kezdeményeinek megjelenését mutatja, gumószeletek Agrobacterium szuszpenzióval történt együtt-tenyésztése után. A rendelkezésre álló kísérleti tapasztalatok szerint egy- és kétszikű fajok esetében egyaránt eredményesen használható. Az Agrobacterium közvetítésével történő transzformáció mellett a plazmidmolekulák közvetlenül is bejuttathatók a növényi sejtekbe. Így a sejtfal enzimatikus eltávolításával létrehozott protoplasztok külső membránja a DNS- molekulák számára átjárhatóvá tehető kémiai és elektromos kezeléssel. A kialakuló pórusokon át a plazmid bekerül a citoplazmába, és ezzel lehetővé válik a sejtmagi integráció. Ez az eljárás igen hatékony, de csak azoknál a növényeknél alkalmazható, amelyek protoplasztjaiból fertilis növények nevelhetők fel. A 2. színes kép kukorica transzformáns növények előállításának lépéseit szemlélteti, a kallusz kolóniák kialakulásától a talajba kiültethető növény felneveléséig. Alternatív megoldást jelent, ha a fém (wolfram) részecskékhez kötött DNS-t belőjük növényi szövetekbe. A sejtmagokat ért találatok esetén a DNS leoldódása után megtörténhet a beépülés. Előnye a közvetlen DNS-beviteli módszernek, hogy a transzformáció előtt a vektormolekulákról eltávolíthatók a nem kívánt szekvenciarészek. Így nem jelent problémát a bakteriális antibiotikumrezisztencia-gén kivágása. Bár kísérleti adatok nincsenek a bakteriális szekvenciáknak a növényi DNS-től eltérő degradációjára a tápanyag lebomlásakor, mégis, ha ezzel megnyugtathatók a fogyasztók, illetve a géntechnológia használatát ellenzők tábora, akkor célszerű élni ezzel az egyszerű megoldással és tanácsos csak növényi szekvenciát hordozó vektort használni a transzformációhoz.
A géntechnológiával nemesített (GN) betegség- és kártevőrezisztens fajták és szerepük a biotermesztésben
Világméretekben igen jelentős, akár 40%-os veszteséget is okozhatnak a különböző kórokozók, amelyek a növények termését, mind a szántóföldön, mind betakarítás után csökkenthetik. Bár a vegyszeres növényvédelem fontos, integráns része a napjainkban alkalmazott növénytermesztési technológiáknak, egyre inkább megfogalmazódik az igény a környezet vegyszerterhelésének mérséklése, illetve az emberi egészséget nem veszélyeztető technológiák használata iránt. Ezekben a törekvésekben kiemelt szerep hárul a rezisztencianemesítésre. Ellenálló fajták használatával válik megalapozottá a biotermesztés. A kémiai növényvédő szerek mellőzése csak akkor érheti el célját, ha egyúttal a növények rezisztenciagénjeik révén képesek a kórokozók elszaporodását mérsékelni. Vajon milyen értéket jelentenek azok a biogabona vagy biotakarmány tételek, amelyek az erős fuzáriumfertőzés következtében nagy mennyiségű toxint tartalmaznak? A fajta-előállítás során a nemesítők állandóan értékelik a különböző tenyészanyagok betegségekkel és kórokozókkal szembeni érzékenységét. Folyamatosan keresik a rezisztenciaforrásokat és keresztezéssel hasznosítják ezeket az értékes géneket, amelyeknek létezését, a genetikai térképen való elhelyezkedését a kísérletek egyértelműen mutatják. A rezisztenciagének szerencsés esetben megtalálhatók a köztermesztésben lévő tenyészanyagokban. Gyakran azonban rokon vadfajokat kell bevonni a rezisztencianemesítésbe. Ezzel jelentősen megnehezül a nemesítő munkája, hiszen keresztezéskor nagy számban kerülnek át rossz, nem kívánt tulajdonságok a kultúrnövénybe. Ezért több generáción keresztül végzett visszakeresztezéssel és állandó szelekcióval lehet csak biztosítani az értékes tulajdonsággal rendelkező növényegyedek felszaporodását. Nehezíti az előrehaladást, hogy egy adott rezisztenciagén a kórokozó változatai közül csak néhány esetében hatásos. Mindig várható új biotípusok megjelenése, amelyek életre szóló elfoglaltságot biztosítanak a nemesítőnek. Az állandó versenyben a nemesítő előnyre tehet szert, ha bővül a hozzáférhető rezisztenciagének köre, illetve növelhető a génkészlet átalakításának precizitása. Így érthető, hogy igen nagy érdeklődést és jelentős kutatási-fejlesztési kapacitást vonz a géntechnológiai megközelítések felhasználása. Az ötletek, a kísérletesen kipróbált stratégiák száma végtelen. Néhányuk használhatóságát az első termesztési tapasztalatok meggyőzően igazolták. Elsősorban a vírusrezisztens és rovarkártevőknek ellenálló fajták elterjedését mutatják a statisztikák. Érdemes megemlíteni, hogy a legsikeresebb megoldások alapötlete jól ismert a korábbi kórtani kutatásokból. Így a kutatók már régen megfigyelték a keresztvédettség jelenségét, amikor valamilyen vírussal fertőzött növény védett a további újrafertőzésekkel szemben. A fertőzött sejtekben jelenlévő virális eredetű molekulák védő hatását a transzgenikus növényekkel végzett kísérletek kristálytisztán igazolták. Ugyanis akár vírus burokfehérjének, akár RNS- molekularésznek a génjeit fejeztették ki transzformánsokban, vírus- ellenállóságot lehetett kimutatni. Szintén régi növényvédelmi gyakorlatra épül a rovarrezisztens transzformáns növények előállításának koncepciója. Ismert, hogy a Bacillus thuringiensis (Bt) szuszpenzióval végzett permetezés több rovar esetében is jó védelmet biztosít. A baktérium által termelt endotoxin a rovar bélrendszerében a hámsejtek membránjának károsításával pusztítja el a kártevőt. Maga a toxinfehérje igen specifikusan csak bizonyos rovarokban fejti ki hatását, ezért a biológiai növényvédelem egyik leggyakrabban használt eljárása a B. thuringiensis sejtszuszpenzió kijuttatása. A géntechnológusok ezeket a tapasztalatokat hasznosították, amikor a toxingént növényekbe építették, és a fehérjét a növényekkel termeltették. Bakteriális génről lévén szó, a toxinszintézis hatékonyságát jelentősen lehetett növelni a DNS kémiai szintézisével. A Bt. toxint termelő burgonya, gyapot és kukorica termesztése néhány év alatt jelentősen megnőtt. Ez a kedvező gazdasági és környezetvédelmi hatásoknak köszönhető. A burgonyabogár-rezisztens fajták használata esetén 40%- kal lehetett csökkenteni a rovarölő permetezőszerek mennyiségét. A Bt. szója termesztése 1998-ban elérte a 16 millió hektárt. Az Amerikai Egyesült Államokban a szója vetésterületének 40%-át ilyen fajták foglalták el.
A génizolálási és transzformációs módszerek segítségével jelentősen kibővültek ismereteink a növény-patogén kölcsönhatás molekuláris mechanizmusaival kapcsolatban. Az inkompatibilis (rezisztens) kölcsönhatás során megnyilvánuló védekezési választ, az ún. hiperszenzitív reakciót (HR), először Klement Zoltán írta le. A HR hatására a fertőzés helyén sejthalál következik be, ami gátolja a fertőzés további terjedését. Az inkompatibilis kapcsolatot jelzi még reaktív oxigéngyökök (szuperoxid anion: O-•; hidrogén-peroxid: H2O2) képződése és a védekezési gének gyors aktiválódása. A kórokozó a gazdanövényt "gén-gén" kölcsönhatás során ismeri fel, ami azt jelenti, hogy a baktérium domináns avirulencia (avr) génjének megfelelő szintén domináns rezisztenciagén, (R), működik a növényben. A rezisztenciagének szerepét régen megfigyelték, mégis csak több ilyen gén izolálása után lehetett a fehérjék szerkezetét és funkcióját feltárni. Az utóbbi évek funkcionális genomprogramjainak köszönhetően több R gén termék vált ismertté, mint azt az 5. ábra [10] szemlélteti. Láthatjuk, hogy ezek a fehérjék vagy a növényi sejt külső membránjában vagy annak közelében helyezkednek el. Majd mindegyik tartalmaz leucin aminosavban gazdag szakaszt, amely a fehérje-fehérje kölcsönhatás kialakításában játszhat szerepet.
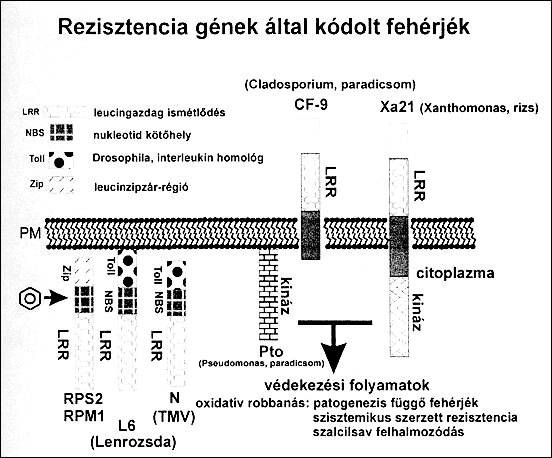
A kórokozók széles körét érintően sikerült izolálni különböző növényekből. A leucin- ismétlődésben gazdag fehérlerégió igen gyakori komponense ezeknek a géntermékeknek. A kinázok fehérjék foszforilációjával vesznek részt a jelátviteli folyamatokban. A membránon (PM) túlnyúló fehérjeszakaszok szerepet játszanak a bakteriális avirulencia (avr) gének termékeivel való kölcsönhatásában. Ez a kölcsönhatás létrejöhet a növényi sejten belül is.
A paradicsom Pto, illetve a rizs Xa21 R gének szerin/treonin kinázokat kódolnak, megerősítve a foszforilációs módosítások jelentőségét a betegségrezisztenciában. Ugyanis több kináz részvételével egy foszforilációs láncolat elindulása aktiválja a sejt védekezési mechanizmusait. Így bekövetkezik a szabadgyökök képződésével az oxidatív robbanás, a programozott sejthalál. A hidrogén-peroxid, H2O2, képződése jelként szolgál a HR reakció elindításához, a védekezési gének bekapcsolásához. A szalicilsav másodlagos jelátvivőként részt vesz széles spektrumú védettséget, az ún. szisztemikusan szerzett rezisztenciát biztosító folyamatok kialakulásában. Ennek során a tünetek mérséklődnek a növény azon részein is, amelyek nem kerültek közvetlen kapcsolatba a kórokozóval. A növényi rezisztenciagének (R) izolálását követően kézenfekvő próbálkozás a transzgenikus növények létrehozása. A rizs Xanthomonas oryzae pv. oryzae-val szembeni ellenállósági Xa21 lokusz genomi DNS-ének felhasználásával előállított rizstranszformánsok 6 rasszal szembeni rezisztenciát mutattak.
A géntechnológiai stratégiák közül kiemelt jelentőséget tulajdoníthatunk azoknak, amelyek a növényekben már működő, természetes védekezési mechanizmusok hatékonyságát növelik. Ilyenkor kevésbé kell számolnunk nem kívánt mellékhatások megjelenésével. Többek között figyelmet érdemelnek a sejteket többszörösen károsító reaktív oxigéngyökök (ROS) képződését befolyásoló molekuláris folyamatok. A növényeket akár biotikus (vírus-, baktérium-, gombafertőzés), akár abiotikus (alacsony vagy magas hőmérséklet, vízhiány, sugárzás és öregedés) stresszhatások érik, az oxigéngyökök létrejötte döntő szerepet játszik a biológiai funkciók károsításában.

A reaktív oxigéngyökök (ROS) képződésének főbb lépései. A sejtek károsításában igen aktív hidroxilgyök (OH•) kialakulásában szerepe van a szabad vas (Fe2+) jelenlétének.
A ROS-képződés fontosabb lépéseit a 6. ábra [11] mutatja be. A szuperoxid anion és a hidrogén-peroxid eltávolítását a sejtekből többféle enzim is elvégezheti. Ebből kiindulva számos kísérletet végeztek szuperoxid-dizmutázt (SOD), katalázt és aszkorbát-peroxidázt kódoló gének túltermeltetésére transzgenikus növényekben. Az oxidatív stresszel szembeni ellenállóság ugyan változott az egyes transzformánsok esetében, azonban többségükben a hatás megbízható volt. A 6. ábrán az is látható, hogy az igen káros hidroxilgyökök (OH•) képződéséhez vas (Fe2+) szükséges. A hidroxilgyökök felelősek a lipidperoxidációért, a fehérje- és DNS- molekulák károsításáért. Mindezek alapján felvetődik a kérdés, lehet-e az oxidatív sejtkárokat mérsékelni a szabad vastartalom csökkentésével. Valamennyi sejtben vaskötő fehérjék, mint pl. a ferritin, szabályozzák a szabad vas mennyiségét. A növényi ferritint kódoló gének működésbe lépnek stressz hatására, illetve abszcizinsav-kezelést követően. Vastúladagolás is aktiválhatja ezeket a géneket a növények vegetatív szöveteiben. A ferritin cDNS-ek izolálásával lehetővé vált ennek a vaskötő fehérjének a folyamatos, magas szintű termeltetése transzformáns dohánynövényekben [12]. A ferritint túltermelő növények kevésbé károsodtak paraquat-kezelés hatására. Ez a korábban gyomirtóként használt szer szabad oxigéngyököket generál a fotoszintetizáló növényi részekben.

A bakteriális, illetve vírusfertőzés okozta tünetek mérsékeltebb formában jelennek meg a ferritint szintetizáló transzformánsok (A2,C8) levelein.
Az oxidatív stresszel szembeni rezisztencia ismeretében feltételezhető, hogy a ferritintranszformánsok megváltozott tüneteket mutatnak kórokozókkal történő fertőzés esetén. A 7. ábrán látható, hogy jelentősen csökkentek a léziós tünetek a Pseudomonas tabaci-val, illetve vírussal történt fertőzést követően. Ezek a transzformánsok nekrotikus gombafertőzés esetén is mérsékelt tünetkialakulást mutattak. A ferritin vaskötő fehérjét túltermelő transzformánsok módot adtak a szabad vasnak az oxidatív károsodásban betöltött szerepének igazolására, illetve új példát szolgáltattak a kórokozókkal szemben toleráns növények előállítására.
GN fajták elterjedése a valós előnyöktől és kockázatoktól függ
Az első géntechnológiai módszerekkel megvalósított génbeépítésről, transzgenikus növényekről szóló közleményeket 1983/84-ben publikálták. Négy évvel később megkezdődött a transzgenikus növények szabadföldi kipróbálása. 1994-ben jelent meg a piacon a géntechnológiával nemesített növényből származó első termék, az etilén anyagcserében megváltoztatott paradicsomból készült püré. Ez a sikeres próbálkozás méreteiben elhanyagolható volt, ugyanakkor megmutatta, hogy megfelelő piacpolitikával (jó minőség, kisebb ár) biztosítható a kedvező vásárlói fogadtatás nemcsak az Amerikai Egyesült Államokban, hanem Angliában is. A ráfordított kutatási-fejlesztési kapacitásokkal arányban, a 90-es évek közepétől már nemcsak évente több ezer szabadföldi kísérletet végeztek, hanem megindult a szántóföldi termesztés is. A géntechnológiával nemesített (GN) fajták vetésterülete évről évre jelentősen nőtt, és így 1999-ben elérte a 40,1 millió hektárt [13].
A statisztika nem teljes, mert valójában nem ismert, hogy például Kínában mekkora területen termesztenek ún. GN fajtákat. Egyes becslések szerint több millió hektárt foglal el csak a rovarrezisztens Bt. gyapot. Igen fontos információt jelent a GN fajták termesztésének földrajzi eloszlása. Az adatok szerint majdnem kizárólag az amerikai kontinensre korlátozódik a jelenlegi hasznosítás [14]. Ezek az arányok igen súlyos megosztottságot tükröznek, aminek különböző okai lehetnek. Talán megismétlődik az információs technológiai fejlesztések idején már tapasztalt európai lemaradás. Az tény, hogy a legfontosabb szabadalmak, az újdonság értékű géntechnológiai eredmények amerikai cégek tulajdonában vannak. Évről évre nő a szakadék, annak ellenére, hogy az EU országai jelentős összegeket fordítanak a kutatásra, segítik a kis vállalatok alapítását ezen a területen. Mindez kevés a lemaradás kedvezőtlen gazdasági és társadalmi következményeinek elhárításához.
Szignifikáns a különbség az európaiak és amerikaiak között a biotechnológia különböző ágazatainak támogatásában
A GN fajták termesztésbe vonását Európában több tényező is késlelteti. A multinacionális cégek gazdasági dominanciája elleni védekezés részét képezi a hasznosítást nehezítő feltételek kialakítása. Nagyon fontos faktor a közvélemény fogadókészsége a GN fajták termékei iránt. Érdekes tanulsággal szolgálnak az európai és amerikai lakosság körében végzett közvélemény-kutatások. A 8. ábra [15] adatai szerint az európaiak egyedül a molekuláris diagnosztikai módszerek használatában mutatnak nagyobb nyitottságot. Véleményük legélesebben a GN fajták termesztése és élelmiszerként történő hasznosítása kérdésében különbözik az amerikaiakétól. Gaskell és mtsai [15] ennek okát három tényezőben látják:
A sajtó befolyása: különösen a lehetséges technológiai ellentmondá-sokkal foglalkozó cikkek számának növekedése segíti az elutasító hozzáállás kialakulását;
Az amerikaiak jobban megbíznak a szabályozásért felelős szervek véleményében, mint az öreg kontinens lakói. Az amerikai szabályozás kiindulópontja, hogy a biotechnológia nem jelent különleges kockázati forrást, és a termékeket a meglévő szabályozási rendszerben kell értékelni. Ezzel szemben Európában a biotechnológiát új eljárásnak tekintik, amihez új szabályzók kellenek;
A tudományos ismeretek színvonala mellett döntő tényező a megfélemlítés mértékében megfigyelhető különbség. Négyszer több európai, mint amerikai gondolja azt, hogy csak a transzgenikus élelmiszerek tartalmaznak DNS-t, géneket vagy azt, hogy a transzgenikus eredetű élelmiszerek fogyasztásával genetikai fertőzés következhet be, illetve minden transzgenikus állat nagyobb. Így nem meglepő, hogy a legalapvetőbb biológiai, genetikai ismeretek hiányában könnyen félrevezethetők, megfélemlíthetők az emberek. Ehhez jó táptalajt nyújt a média szenzációt hajhászó tájékoztatási gyakorlata. A félrevezetés eredményességét segítik a rossz, félbeszakadt kísérletek, az ún. szakértői vélemények, amelyeket nem támasztanak alá kutatási eredmények. A harcos ellenzők táborában sokszor jutnak szóhoz olyan "jól tájékozott" kívülállók, akik saját kísérleti tapasztalatok nélkül, a tudományos kutatómunka szabályait figyelemen kívül hagyó forrásokra hivatkozva fejtik ki elítélő véleményüket a géntechnológiáról.
Természetesen a kialakult európai helyzetért felelősek a géntechnológiai fejlesztéseket végző kutatók, a termékeket előállító és forgalmazó cégek szakemberei is. Nem elég eredményes például a meglévő tudományos ismeretanyag népszerűsítő terjesztése, pedig erre igen nagy szükség van, hogy az emberek a tudományos tények birtokában alkothassanak véleményt. Hazánkban az egyetemi tananyagok magas színvonalon tartalmazzák a molekuláris biológiára épülő géntechnológiai ismereteket. Ugyanakkor kívánatos lenne a gimnáziumi és általános iskolai biológiatanítás anyagába beépíteni a legfontosabb új információkat. A biotechnológiai, agrokémiai cégek több súlyos hibával járultak hozzá a kedvezőtlen vásárlói fogadtatás kialakulásához. Az egyik, hogy nem fektettek megfelelő hangsúlyt a technológia alapjainak, a termékek tulajdonságainak bemutatására. A másik: fejlesztési stratégiájuk kidolgozásakor nem a vásárlót közvetlenül érintő tulajdonságok javítása került előtérbe. Nem éppen szerencsés érvelés, hogy a termesztésben lévő GN fajták 70%-a gyomirtó szerekkel szemben rezisztenciával rendelkezik. Bár ezek a tenyészanyagok kedvezőbb technológiai hátteret nyújtanak a gazdaember számára, mégis a termék fogyasztóját választásában nem befolyásolják a növénytermesztési szempontok. A GN fajták 22%-a rovar- és vírusrezisztencia géneket hordoz. Ezek a kedvező tulajdonságok is még elsősorban az agrotechnika javítását szolgálják, ugyanakkor ebben az esetben igen jelentősek a környezeti és egészségvédelmi pozitívumok. A kémiai növényvédő szerek használatának mérséklése már lehet olyan többlet, amely irányítja a vásárlói szokásokat. Mégis a géntechnológiai termékek számottevő elterjedését akkor várhatjuk, ha ezek az áruk olcsóbbak és jobb minőségűek lesznek, ha garantálják az egészséget veszélyeztető összetevők (akár szermaradványok, akár a beépített gén működésével összefüggő anyagok) hiányát a termékben. Nem nehéz megjósolni, hogy az ezeknek a kritériumoknak megfelelő termékek fejlesztése világszerte egyre jelentősebb kapacitással folyik.
Érdemes felhívni a figyelmet azokra a különbségekre, amelyek a géntechnológia hatásait illetően a fejlődő és fejlett államokban fellelhetők. A mezőgazdasági túltermelés gondjaival elfoglalt országok esetében érthető az elzárkózás minden kezdeményezéstől, amely többlettermést, újabb problémákat okozhat. Elsősorban az élelmiszerek minősége, illetve a megszokott tradíciók irányítják a vásárlói szokásokat ezekben a fejlett országokban. Ugyanakkor a Föld élelmiszerhiánnyal küzdő részein minden technológia, amely többlettermést eredményez, igen nagy jelentőségű. Különösen a kedvezőtlen adottságú területeken folytatott növénytermesztés igényli a biológiai potenciál javítását, a növények életfolyamatainak optimalizálását. Egy-egy betegség-ellenálló vagy szárazság-, sótűrő tenyészanyag sokat segíthet embermilliók szenvedésének mérséklésében. Mivel a kedvező hatású gének a helyi tájfajtákba építhetők be, így biztosítható a meglévő mezőgazdasági kultúrák fenntartása, és az eredményesség javítása. A géntechnológia alkalmazásában ezek az országok rászorulnak a fejlett világ támogatására. Nemzetközi szervezetek kiemelt szerepvállalása mellett a biotechnológiai cégek nyitottságára is szükség van. Természetesen a fejlődő országok saját kutatási-fejlesztési tevékenysége is meghatározó jelentőségű. Így például Kína már kiterjedten használja a géntechnológiával nemesített fajták előnyeit. A jövő ad választ arra a kérdésre, hogy milyen szerepet játszanak majd ezek a tenyészanyagok 20-40 év múlva, amikor - az előrejelzések szerint - a mainál kétszer több élelmiszert kell termelni a Föld megnövekedett népessége számára.
Az elkövetkező években a géntechnológia szerepvállalását jelentősen befolyásolja majd a törekvés, hogy megfeleljen a jogos elvárásnak: azaz, hogy a géntechnológiával nemesített növények termékei a lehető legkisebb kockázatot jelentsék a fogyasztó egészsége és a környezete számára. Számos félreértésre vagy tudatos félremagyarázásra adhat lehetőséget a felületes általánosítás. Nem lehet általában azt állítani, hogy a kalapos gombák veszélyt jelentenek, hiszen nap mint nap fogyasztunk ízletes, ehető gombákat. Minden egyes transzgenikus növény esetében növény- és génkombinációnként kell értékelni a kockázat mértékét. Így más megítélés alá esik például, ha növényi szekvenciákat építenek be. Lényeges kérdés, hogy a beépített gén terméke jelen van-e a termékben, és milyen biológiai hatásai lehetnek. Magától értetődő követelmény, hogy amennyiben új hatóanyag jelenik meg a fogyasztott termékben, akkor kiterjedt, az élelmiszer-minősítés előírásainak megfelelő módon el kell végezni a szükséges toxikológiai, mutagénteszteket. Ezzel egy időben túlzás egy vírus- vagy herbicidrezisztens cukorrépából készült cukor részletes vizsgálatát megkövetelni. A törvényi szabályozás lehetőséget ad a mérlegelésre. Talán joggal feltételezhető, hogy a termék előállítójának jól felfogott érdeke a minőség biztosítása. Az árunak a paraméterei fontosak és nem az, hogy milyen módszerrel - keresztezéssel, mutációval vagy poliploidizációval - állították elő. Könnyen létrehozható keresztezéses nemesítéssel olyan tenyészanyag, amely nem kívánt hatóanyagokat tartalmaz. Ezek kiszűrését a növénynemesítő, a fajtaminősítés és élelmiszer-ellenőrzés szakemberei hivatottak elvégezni. Mivel a transzgenikus genotípusok is bekerülnek az értékelés hasonló folyamatába, több generáción át végzett minősítés garantálhatja a nem kívánt változatok eltávolítását és csak az értékes növények további felhasználását.
A GN fajták használatának ellenzői veszélyeztetve látják a természetes növénypopulációkat, hiszen számos vadon élő faj kereszteződhet a kultúrnövényekkel. Ez egy fontos, figyelmet érdemlő probléma, ezért indokolt kísérletesen is megvizsgálni a transzgén átjutásának esélyeit a vadon élő fajokba. Kiterjedt kísérletezési tapasztalat mutatja, hogy számos termesztett növényünknek (pl. búza, kukorica, burgonya stb.) nincs olyan rokon faja, amellyel spontán kereszteződne. Így ezekben a fajtákban a transzgén biztonságban van. A lucerna vagy repce esetében a beépített gének átkerülnek gyomnövényekbe, így ezekben az esetekben körültekintően kell a hatásokat értékelni. Megfontolandó annak végiggondolása, hogy ilyenkor a transzgén ugyanolyan valószínűséggel kereszteződik át, mint a kultúrnövény többi génje. Tehát a vad fajok ilyen úton történő nemesedése már hosszú idő óta bekövetkezhetett volna, hiszen a termesztett növény számos génje jelenthet szelekciós előnyt egy gyom számára. A tapasztalat azt mutatja, hogy a gyomok populációi nem változnak a kultúrnövény irányába. Sajnos az ellenkező irányú génáramlás jeleit sem látjuk, hogy a szárazságtűrő gyomok génjei spontán módon javítanák a fajták stressz-rezisztenciáját.
A biológiai sokféleséget is sokan féltik a transzgenikus növény elterjedése kapcsán. Ebben a kérdésben az álláspontok közelítését nehezíti, hogy nehéz kísérletet tervezni a probléma vizsgálatára. Abból kellene kiindulni, hogy függetlenül a GN fajták használatától, az évszázadok során kialakult növénytermesztési gyakorlat szüntette meg a szántóföldeken a biológiai sokféleséget. A fajtaelőállítás elsődleges szempontja, hogy egyetlen faj igen hasonló egyedeit neveljük, ami lehetővé teszi a gépesítést és a hatékony földhasználatot. Szántóföldjeinken a fajok sokféleségét minden évben gyomirtással szüntetjük meg. A termesztett növények populációi alapjaiban térnek el a természetes ökoszisztémáktól. Nemesítési szempontból a transzgenikus növények előállítása növeli a genetikai variabilitást. Ennek a technológiának éppen az az előnye, hogy a specifikus hatást a genotípusok széles körét érintően el lehet érni. Nincs hasonló nemesítési módszer, amely egy-két év leforgása alatt több száz fajtában ki tudna alakítani egy új tulajdonságot. Ma már nagy fajtaválasztékot kínálnak a géntechnológia módszerét használó nemesítő cégek.
Látva az európai és amerikai stratégiák különbözőségét a géntechnológiai fejlesztések területén, igen jelentős kihívás Magyarország számára a lehetséges mozgásterület megtalálása. Érdemes abból kiindulni, hogy hazánk klimatikus és talajadottságai jobbak, mint az európai és világátlag. Igen jelentős növénynemesítési hagyományokkal rendelkezünk. A magyar nemesítők fajtái sokszor piacra találnak külföldön is. Meghatározó szerepű a vetőmagipar. A nemesítés és vetőmagtermesztés olyan terméket eredményez, amelyben jelentős hozzáadott szellemi érték testesül meg. A fentiek alapján elsőrendű érdekünk ennek a tevékenységnek a versenyképességét biztosítani. Ha hihetünk a bemutatott kutatási eredményeknek és az amerikai kontinensen tapasztalható fejlesztések gazdasági fontosságának, akkor érdemes felkészülni arra, hogy élni tudjunk a géntechnológia által nyújtott előnyökkel. A kutatás oldaláról az alapfeltételek megteremtése ideje korán megkezdődött, hiszen 1986-ban, 2-3 évvel az első transzgenikus növények közlése után megjelent Deák és mtsai cikke [16] a lucernába történt génbeépítésről. Azóta a magyar növénynemesítők közreműködésével kukorica, repce, burgonya, dohány, rizs, búza transzgenikus növényeket állítottak elő idehaza. Több agronómiai szempontból jelentős gén izolálásáról is beszámoltak kutatóink. Különösen az EU-csatlakozás előtti felkészülés időszakában lehet indokolt, hogy hatékonyan kihasználjuk a meglévő szellemi és metodikai alapokat, hiszen így lehet kedvező versenyhelyzetet biztosítani és felkészülni a jövő kihívásaira.
A kedvező amerikai tapasztalatokat alapul véve és feltételezve, hogy a közvélemény előbb vagy utóbb meggyőződik a géntechnológiai termékek előnyeiről és megbízhatóságáról, jelentős fejlődést és kiélezett versenyt prognosztizálhatunk. Ez a technológia jelenlegi kezdeti formájában még további igen intenzív kutatómunkát és tökéletesítést igényel. Mindez nem jelentheti azt, hogy elutasítjuk ezt a tudomány által felkínált lehetőséget, hiszen folyamatosan több és jobb minőségű élelmiszerre van szükség ezen a bolygón.
Köszönetnyilvánítás Ezúton szeretnék köszönetet mondani Keczán Józsefné Czakó Zsuzsannának a kézirat gépelésében és az ábrák elkészítésében nyújtott segítségéért.
IRODALOM
1. Koncz, Cs., Sümegi, J., Udvardy, A., Ramcsány, M. and Dudits, D. (1981) Cloning of mtDNA fragments homologous to mithochondrial S2 plasmid-like DNA in maize. Mol. Gen. Genet., 183: 449-458.
2. Dudits, D. és Heszky, L. (1990) Növény- biotechnológia, Mezőgazdasági Kiadó, 310 oldal.
3. Dudits, D. és Heszky, L. (2000) Növényi biotechnológia és géntechnológia, Agroinform Kiadóház, 300 o.
4. Balázs, E. és Dudits, D. (szerk.) (2000) Szemelvények a molekuláris növénybiológiából Akadémiai Kiadó,
5. Lin X., Kaul, S., Rounsley, S., Shea, T.P., Benito, M-I., Town, C.D., Fujii, C.Y., Mason, T., Bowman, C.L., Barnstead, M., Feldblyum, T.V., Buell, C.R., Ketchum, K.A., Lee, J., Ronning, C.M., Koo, HL., Moffat, K.S., Cronin, L.A., Shen, M., Pai, G., Van Aken, S., Umayam, L., Tallon, L.J., Gill, J.E., Adams, M.D., Carrera, A.J., Creasy, T.H., Goodman, H.M., Somerville, C.R., Copenhaver, G.P., Preuss, D., Nierman, W.C., White, O., Eisen, J.A., Salzberg, S.L., Fraser, C.M. and Venter, C. (1999) Sequence and analysis of chromosome 2 of the plant Arabidoposis thaliana. Nature, 402: 761- 768.
6. Mayer, K. et al. (1999) Sequence and analysis of chromosome 4 of the plant Arabidopsis thaliana. Nature, 402: 769-777.
7. Benfey, P.N. Chua, N.H. (1990) The cauliflower mosaic virus 35S promoter: combinatorial regulation of transcription in plants. Science, 250: 959-966.
8. Ho, M-W., Ryan, A. and Cummins, J. (1999) Cauliflower mosaic viral promoter - A recipe for disaster? Microbial Ecology in Health and Disease, 11: www.scup.no/mehd/ho
9. Hammond, B.G., Vicini, Jéki László, Hartnell, G.F., Naylor, M.W., Knight, C.D., Robinson, E.H., Fuchs, R.L. and Padgette, S.R. (1996) The feeding value of soybeans fed to rats, chickens, catfish and dairy cattle is not altered by genetic incorporation of glyphosphate tolerance. American Institute of Nutrition, 717-727.
10. Baker, B., Zambryski, P., Staskawicz, B. Dinesh- Kumar, S.P. (1997) Signaling in plant-microbe interactions. Science, 276: 726- 733.
11. Hammond-Kosack, K. E. Jones, J.D.G. (1996) Resistance gene-dependent plant defense responses. The Plant Cell, 8. 1773--1791.
12. Deák, M., Horváth, V.G., Davletova, S., Török, K., Sass, L., Vass, I., Barna, B. Király, Z. and Dudits, D. (1999) Plants expressing the iron-binding protein, ferritin ectopically are tolerant to oxidative damage and pathogens. Nature Biotechnology, 17: 192-196.
13. Ferber, D. (1999) GM crops in the cross hairs. Science, 286: 1662-1666.
14. Serageldin, I. (1999) Biotechnology and food security in the 21st century. Plant Biotechnology: Food and Feed, 387- 389.
15. Gaskell, G., Bauer, M.W., Durant, J., Allum, N.C. (1999) Worlds apart? The reception of genetically modified foods in Europe and the U.S. Plant Biotechnology: Food and Feed, 384-387.
16. Deák, M., Kiss, Gy.B., Koncz, Cs. and Dudits, D. (1986) Transformation of Medicago by Agrobacterium mediated gene transfer. Plant Cell Reports, 5: 97-100.
SZÍNES KÉPEK FELIRATA
1. kép A lucerna hisztongén rövid DNS- szakasza merisztéma specifikus génkifejeződést biztosít transzgénikus dohánynövényekben.
A. Egy transzformációs vektor, amely tartalmazza a promoterrégiót és a hozzá kapcsolt (-glukoronidáz (GUS) ripotergént.
B-E. A GUS riportergén működése kék indigófesték felhalmozódását eredményezi, ami jelzi azokat a szövetrészeket, ahol a gén aktív állapotban van.
2. kép Transzgenikus burgonya és kukorica előállításának lépései.
A. Burgonyagumó szeletek fertőzése Agrobacteriummal beépített gént hordozó hajtások differenciálódását eredményezi.
B. Kukorica protoplasztokba történt DNS- molekula bevitele után kiszelektálhatók a transzfrománs kolóniák.
C-D. Hajtáskezdemények differenciálódása transzformáns szövetekből.
E. Vírus burokfehérjegént hordozó kukoricanövényke nevelése táptalajon.
F. Transzformáns kukoricanövény talajba történt kiültetés után.